Dra. Paula Rezende Teixeira
Introdução
As interações celulares são fundamentais para o desenvolvimento e para o bom funcionamento dos organismos. Estão presentes em diferentes momentos da embriogênese e ocupam importantes funções em alguns órgãos e tecidos. Foram descritas pela primeira vez em meados da década de 40, embora pouco se soubesse sobre as proteínas e as interações envolvidas. Experimentos clássicos com células de embriões de anfíbios mostraram a alta seletividade destas interações, permitindo que células similares mantenham-se unidas. Essas adesões seletivas permitiram que as células dissociadas em células individuais e misturadas, puderam se reorganizar como em um embrião inicial, seguindo o padrão onde as células da placa neural se localizaram mais internamente para dar origem ao tubo neural, as células da ectoderme se localizaram mais externamente e entre estas as células mesodérmicas (Figura 1). Com o tempo novas descobertas foram feitas na área e logo a proteína caderina foi identificada como sendo a responsável por essa seletividade específica.

Figura 1 – Movimentos e interações celulares para formação da placa neural. (Créditos: Fábio Siviero)
Atualmente uma série de moléculas de adesão celular são conhecidas, divididas em diferentes classes, assim como diferentes tipos de junções celulares desempenhando diferentes funções. Assim, esse módulo tem por objetivo descrever as principais moléculas de adesão, identificar os tipos de junções que pode ocorrer entre duas células e fornecer alguns exemplos destas interações célula-célula, tanto interações célula-célula estáveis que estão presentes na organização das células em tecidos, quanto interações célula-célula transientes que estão presentes na resposta imune.
Iniciaremos com as moléculas de adesão, que podem ser qualificadas em duas classes principais: 1) moléculas de adesão cálcio (Ca2+) dependentes, sendo: caderinas e selectinas, onde estas moléculas necessitam da presença de cálcio para uma correta interação célula-célula. 2) moléculas de adesão cálcio (Ca2+) independentes, sendo: integrinas e as proteínas da superfamília das imunoglobulinas (CAMs) que não necessitam de cálcio.
Moléculas de adesão
Caderinas
A molécula de adesão caderina pertence a uma superfamília de proteínas, são conhecidas mais de quarenta tipos de caderinas, algumas delas e a onde são expressas estão identificadas abaixo, por exemplo:
- E-caderina, expressa em diferentes epitélios;
- N-caderina, expressa na ectoderme neural e nas células do músculo cardíaco;
- P-caderina, expressa em trofoblastos e pulmão;
- M-caderina, encontrada em mioblastos e músculo esquelético;
- R-caderina, encontrada nas células da glia.
Estruturalmente, essa molécula possui 4 sítios extracelulares onde se ligam cálcio, e ainda na porção extracelular é possível identificar o domínio HVA, representado por três aminoácidos: histidina, valina e alanina; esse domínio é responsável pela dimerização da caderina e posterior interação com outra molécula na célula vizinha. Essas moléculas fazem interações cis-homofílicas, ou seja, interagem com uma molécula idêntica formando dímeros e posteriormente fazem interações trans-homofílicas interagindo com dímeros da mesma molécula na célula vizinha.
No seu domínio citoplasmático, a proteína caderina se liga a actina, no entanto, essa ligação se faz por proteínas de ancoragem. Um complexo de cateninas α, β e γ e proteínas de ligação alfa-actinina, vinculina e formina fazem a ponte entre caderina e actina (Figura 2). Esse complexo de proteínas que liga actina a caderina desempenham papeis importantes, não só ligando actina, mas também regulam a interação e controlam a adesividade extracelular. Essa associação é também importante para morfogênese celular, estabilidade celular e polaridade.

Figura 2 – Ilustração evidenciando a adesão entre duas células, por meio de complexos proteicos e a caderina (amarela). (Créditos: Edilson Jacob)
Figura 3 – Em elaboração.

As caderinas formam junções estáveis e específicas, e é a porção extracelular que responde pela grande variedade de caderinas. Nas primeiras divisões celulares do embrião, quando ainda está sofrendo clivagens, é possível observar a presença das proteínas caderinas, já no embrião de apenas 8 células, onde a caderina é a proteína de adesão responsável pela compactação celular do embrião, formação da mórula (Figura 3). As caderinas controlam a organização seletiva das células, durante o desenvolvimento, a migração ou a separação das células também é dependente do tipo de caderina, mudanças no padrão de expressão de caderinas durante a formação do sistema nervoso levam a uma segregação de grupos de células que expressam determinadas caderinas. As junções também podem ser feita pelos diferentes graus de expressão de uma mesma caderina, células que expressam altos níveis se agrupam e se separam das células que expressam baixos níveis de caderina (Figura 4).

Figura 4 – Ilustração evidenciando como as células epiteliais se separam ou se agrupam com base na expressão de diferentes tipos de caderinas ou níveis de expressão de uma mesma caderina. As caderinas são proteínas de adesão célula-célula dependentes de cálcio. (Créditos: Fábio Siviero)
Selectinas
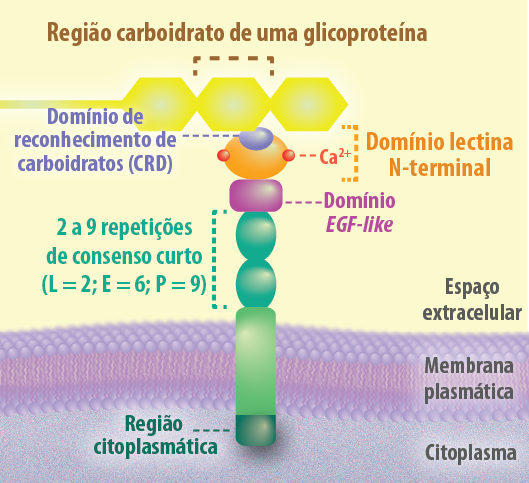
As proteínas selectinas são o segundo grupo de moléculas dependentes de cálcio. No seu domínio extracelular, ligam carboidratos que pertencem ao grupo das lectinas. Cada selectina tem um domínio CRD, com afinidade de ligação a um oligossacarídeo específico aderido a uma proteína (glicoproteína) ou um lipídeo (glicolipídeo). Na porção extracelular é possível encontrar além do domínio de reconhecimento de carboidratos (CRD), específico para um açúcar em particular (galactose, manose, N-acetilglicosamina etc.); um domínio homólogo a uma sequência de repetição encontrada no fator de crescimento epidérmico (EGF-like), e muitas sequências de repetição consensos encontradas em proteínas reguladoras do complemento (Figura 5).
Três principais tipos de selectinas modulam as interações transientes entre leucócitos e células endoteliais ou plaquetas sanguíneas. As selectinas reconhecem carboidratos específicos na superfície de células e medeiam a interação inicial entre leucócitos e células endoteliais durante a migração dos leucócitos para os locais de inflamação, sendo elas:
- L-selectina, expressa na superfície de linfócitos e possui alta afinidade de ligação para carboidratos sulfatados;
- E-selectinas, expressas na superfície de células endoteliais ativadas;
- P-selectina, expressa na superfície de plaquetas e células endoteliais ativadas.
Selectinas, juntamente com integrinas e moléculas de adesão intercelular (ICAMs), desempenham um papel significativo em inflamações e na migração periódica de linfócitos da circulação para órgãos linfoides, processo denominado endereçamento ou homing, que será descrito em detalhe posteriormente.
Figura 5 – Estrutura de uma proteína transmembrana pertencente à família das selectinas, que reconhecem sequências de carboidratos em glicoproteínas e desempenham um papel crucial nas interações célula-célula de curta duração, especialmente no contexto da inflamação e migração periódica de linfócitos. (Créditos: Edilson Jacob)
Superfamília de imunoglobulinas
As moléculas de adesão pertencentes a superfamília das imunoglobulinas são cálcio independentes e codificadas por um único gene. Membros da superfamília são gerados por processamento alternativo (splicing) do mRNA e apresentam diferenças em relação a sua glicosilação. Possuem um segmento extracelular pregueado com 2 a 6 domínios semelhantes a imunoglobulinas, por isso assim identificadas. Podem se ligar a moléculas idênticas em outra célula (ligações trans-homofílicas) ou a outros membros da família das imunoglobulinas, assim como a proteínas diferentes, como integrinas (ligações trans-heterofílicas). São responsáveis, juntamente com as proteínas integrinas pela formação de uma adesão mais estável entre a superfície do leucócito e a superfície endotelial, envolvendo integrinas da superfície dos leucócitos e moléculas de adesão intercelular da superfamília das imunoglobulinas expressas na superfície do endotélio. A adesão firme permite que o leucócito penetre as paredes dos capilares e entrem no tecido adjacente através da migração entre as células epiteliais. Isso resultará na penetração do leucócito em direção ao tecido onde se encontra a inflamação ou injuria.
Integrinas
As moléculas de adesão integrinas são cálcio independentes, e é a única molécula de adesão formadas por um heterodímero com duas subunidades associadas α e β provenientes de genes diferentes. São conhecidos pelo menos 24 heterodímeros de integrinas. A subunidade alfa tem duas cadeias unidas por pontes dissulfeto e uma cabeça globular com sítios para ligação de cátions divalentes. A subunidade beta possui duas características significativas, a cadeia externa contem regiões repetidas ricas em cisteína e a porção intracelular interage com filamentos de actina através de proteínas de conexão: talina, vinculina e α-actinina. Todas as células expressam uma ou mais integrinas, esta molécula de adesão apresenta uma pequena cauda carboxi-terminal intracelular e um grande domínio N-terminal extracelular, onde o domínio extracelular liga-se a sequência tripeptídica RGD (arginina, glicina, aspartato) presente em proteínas da membrana basal, laminina e fibronectina. Integrinas desempenham dupla função, ligam-se a matriz extracelular e ao citoesqueleto no interior da célula (Figura 6). Dessa forma podem estar relacionadas tanto com a migração celular na embriogênese pela ligação com a matriz extracelular, como também participam da interação célula-célula pela interação do heterodímero α1 β2 expresso em leucócitos com ligantes nas superfícies das células endoteliais (interação célula-célula transiente) durante a fase de extravasamento do endereçamento.
Figura 6 – Idem a figura 5 (Créditos: Edilson Jacob)
Junções Celulares
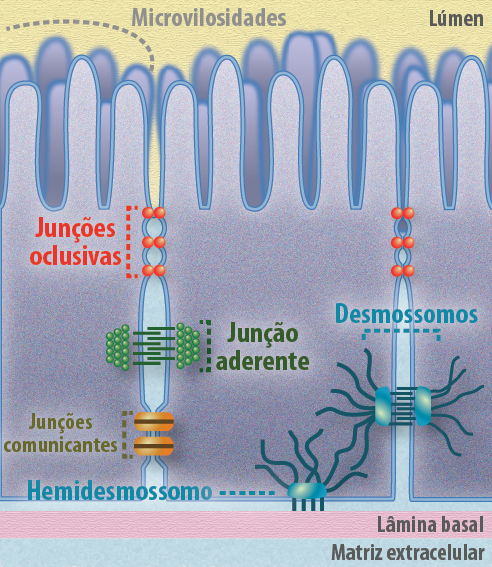
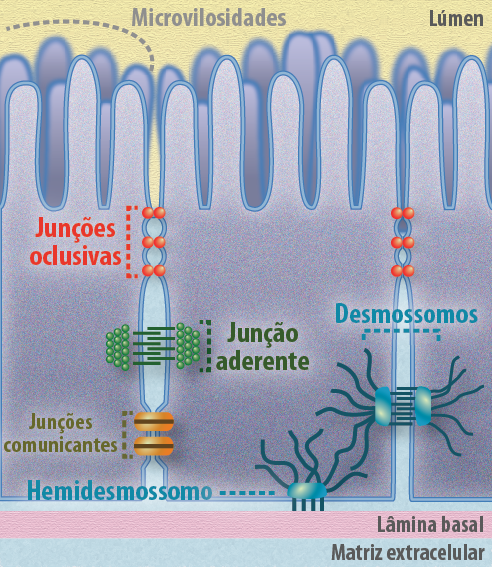
As junções celulares são ligações físicas que respondem pela adesão do tipo célula-célula e fornecem estabilidade e polaridade celular. O movimento de solutos, íons e água em uma camada epitelial pode ocorrer através ou por entre as células. Sendo a via transcelular aquela onde participam uma série de proteínas transportadoras que levam esses solutos para o interior da célula. E a via paracelular aquela onde as junções serão as responsáveis por permitir ou não a passagem desses solutos por entre as células. Existem três classes principais de junções (Figura 7):
- Junções de Oclusão, que selam os espaços entre as células do epitélio, tornando-o uma barreira impermeável ou seletivamente permeável;
- Junções de Ancoragem (zona de adesão e desmossomos), que transmitem o estresse e estão imbricadas aos filamentos do citoesqueleto;
- Junções Comunicantes, que criam passagens ligando citoplasmas de células adjacentes.
Figura 7 – A imagem ilustra os tipos de junções intercelulares, encontradas principalmente nas células epiteliais, essenciais para a coesão do tecido e para a comunicação. (Créditos: Edilson Jacob)
Junções de oclusão
Junções de oclusão possuem duas principais características: determinam a polaridade das células epiteliais e impedem a livre passagem de substancias; representam uma barreira a via paracelular. Duas principais proteínas estão presentes nessas junções claudina e ocludina, que pertencem à família das tetraspaninas com quatro domínios transmembrânicos, constituem um lacre de fechamento do epitélio. A proteína ocludina interage com quatro proteínas sendo: as proteínas da zônula de oclusão ZO-1, ZO-2 e ZO-3 e afadina. As proteínas claudinas formam fibrilas lineares nas junções, é a proteína que de fato sela a passagem via paracelular. Pertence a uma família de 16 proteínas, que serão responsáveis pelas diferentes permeabilidades ou impermeabilidades presentes nos diferentes epitélios. Não formam adesões fortes e por isso o contato é reforçado com as proteínas das zônula de oclusão (ZO-1, ZO-2 e ZO-3) e também afadina. Outras proteínas também se juntam a essa junção, sendo membros da superfamília das imunoglobulinas: nectina e JAM (molécula de adesão juncional). Nectina e JAM se associam a ZO-1 formando cis-homodímeros e em seguida, trans-homodímeros através do espaço extracelular, nectina também se liga aos filamentos de actina através de afadina (Figura 8).
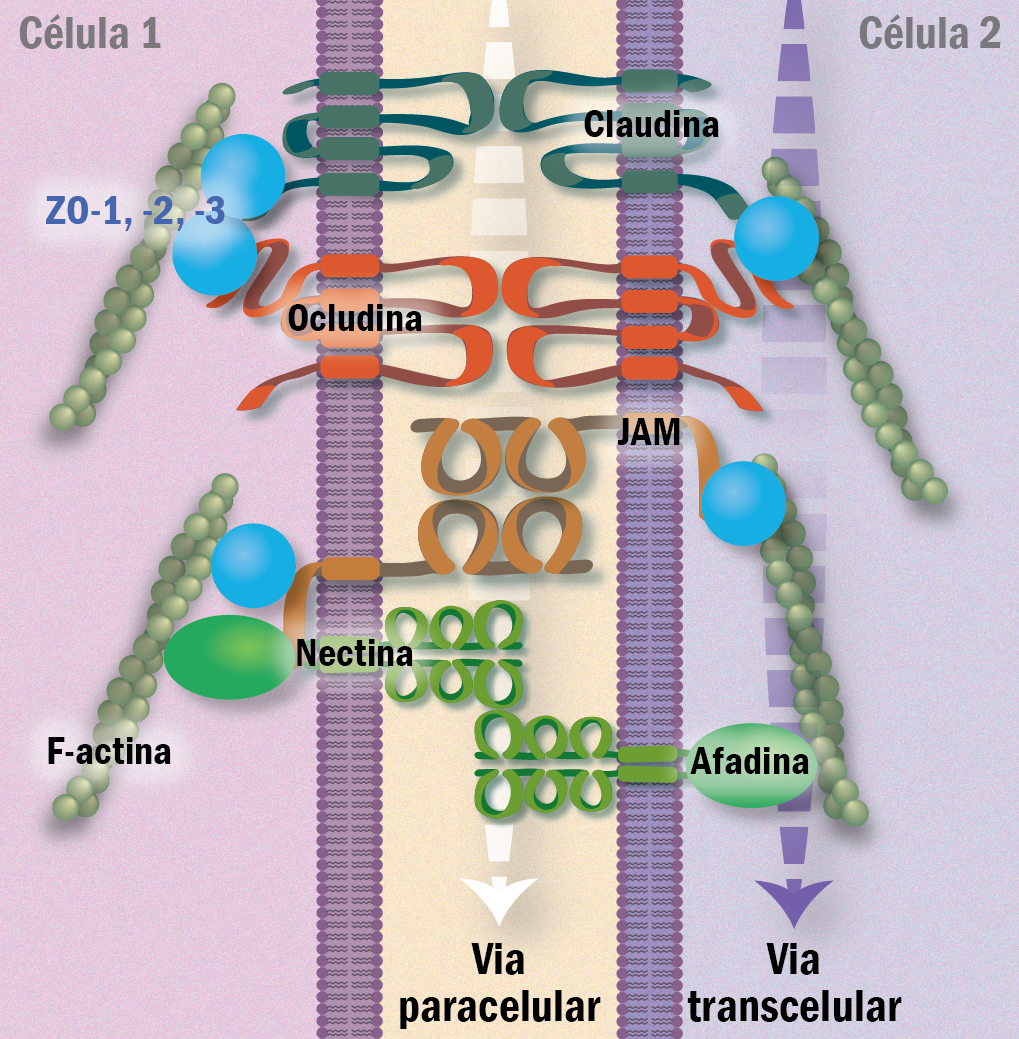
Figura 8 – Estrutura de uma junção oclusiva (tight junction), um tipo de junção célula-célula crucial para criar uma barreira de vedação em epitélios, impedindo a passagem de substâncias entre as células. (Créditos: Edilson Jacob)
As junções de oclusão, também conhecida como zônula de oclusão, estão localizadas na região mais próxima da superfície apical da célula, cobrem todo o perímetro celular semelhante a um cinturão envolvendo toda a circunferência apical da célula. A zônula de oclusão na membrana celular quando analisada por criofraturas ao microscópio eletrônico mostra um aspecto semelhante a um “acolchoado” de faixas anastomosadas (Figura 9). Dependendo do número e dos padrões das faixas na zônula essas junções podem ser permeáveis ou impermeáveis.
Figura 9 – Em elaboração.
Junções de ancoragem
As junções de ancoragem são formadas pela zônula de adesão e pelos desmossomos, que juntamente com a zônula de oclusão formam o complexo unitivo da célula.
A zônula de adesão localiza-se abaixo da zônula de oclusão e também envolvem todo o perímetro apical da célula. São formadas principalmente pela proteína caderina que interage com as proteínas de ancoragem: catenina, vinculina e α-actinina, esse tipo de junção une duas membranas celulares de células diferentes e também promove a associação do citoesqueleto das duas células através das proteínas transmembrânicas de ligação. Essas junções transmitem o estresse gerado pelo citoesqueleto de actina entre as células que estão unidas, essa rede de actina é potencialmente contrátil e proporciona à camada epitelial a capacidade de desenvolver tensão e mudar sua forma de maneira extraordinária, por exemplo: a invaginação da camada epitelial causada por uma constrição organizada ao longo do cinturão de adesão em regiões selecionadas do epitélio levam a importantes modificações morfológicas. O espaço intercelular de 15-20 nm entre os folhetos externos de duas membranas adjacentes é ocupado pelos domínios extracelulares das proteínas caderinas (figura 10). Esse espaço entre as membranas celulares é bem diferente para as junções de oclusão, que chegam ate 5nm, aproximando ainda mais as membranas, garantindo um selamento entre os folhetos.
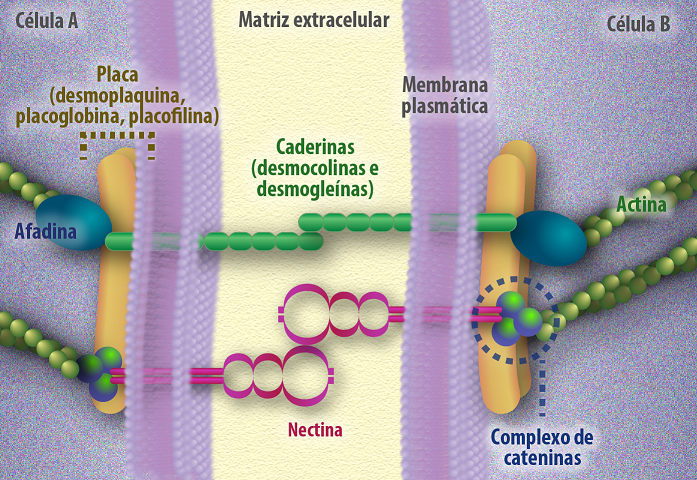
Figura 10 – (Créditos: Edilson Jacob)
Os desmossomos também constituem as junções de ancoragem e diferente da zônula de adesão, estão aleatoriamente distribuídos ao longo da membrana, semelhantes a um ponto de solda, ou seja, não estão circundando toda a célula. Os desmossomos são formados por duas placas de adesão em formato de disco que possuem aspecto bastante elétron-denso ao ME e projeções de 400x2500x10nm, localizadas opostas uma a outra nas faces citoplasmáticas da membrana. Essa placa é formada pelas proteínas da família das caderinas: desmoplaquinas e placoglobinas que interagem com as proteínas, também da família das caderinas, desmogleinas e desmocolinas, localizadas no espaço extracelular. O espaço intercelular chega a 30nm sendo ocupada pela porção glicosilada destas proteínas. A placa de ancoragem liga-se aos filamentos intermediários da célula, sendo filamentos de queratina para junções presentes nas células epiteliais ou filamentos de desmina para junções presentes nas células do músculo cardíaco (figura 11). Essas junções são dependentes de cálcio e sua ausência pode causar quebra nessas junções que podem gerar injurias muito rapidamente. Estas junções respondem pela resistência e rigidez do tecido.
Figura 11 – (Créditos: Edilson Jacob)

Junções comunicantes
Junções comunicantes medeiam a comunicação intercelular por permitir a passagem de várias pequenas moléculas entre citoplasma adjacentes. O canal permite a passagem de moléculas até 1,5kDa pelos poros formados. Íons, açúcares, aminoácidos, nucleotídeos, segundo mensageiros como AMP cíclico ou GMP cíclico entre outras pequenas moléculas atravessam a fenda formada nessa junção, uma fenda estreita e constante com 2-4 nm de espessura. Esses canais são formados pela proteína denominada conexina, são mais de 20 tipos de conexinas conhecidas. As conexinas são proteínas transmembranas que se agrupam para formar o canal, denominado conexon, constituído por 6 proteínas conexinas, que se associam intimamente entre si e com a membrana através dos seus 4 domínios transmembrânicos.
Os conexons podem ser classificados de acordo com a composição das proteínas conexinas que o forma, se as conexinas forem iguais formarão um conexon denominado homomérico, e se forem diferentes conexinas formaram um conexon denominado heteromérico. Um conexon, também conhecido como hemicanal, está emparelhado com seu par na membrana plasmática adjacente, os dois conexon se fundem, formando um canal hidrofílico funcional de comunicação intercelular. Essa interação pode ser do tipo homotípica quando os conexons apresentam a mesma configuração protéica ou heterotípico quando a configuração é diferente entre as proteínas conexinas em cada conexon (Figura 12). Os diferentes tipos de conexons formados podem ter diferentes funções em diferentes tipos celulares, a permeabilidade também é diferente e os canais podem se abrir ou fechar rapidamente dependendo do estado fisiológico da célula. Em condições de pH citossólico baixo e concentração de cálcio alta, os canais se fecham, para manter integridade nas outras células, uma vez que a alta concentração do íons cálcio pode ser conseqüência de dados na membrana plasmática. Na situação inversa, aumento do pH citossólico e diminuição da concentração de cálcio, os canais se abrem permitindo a comunicação celular.
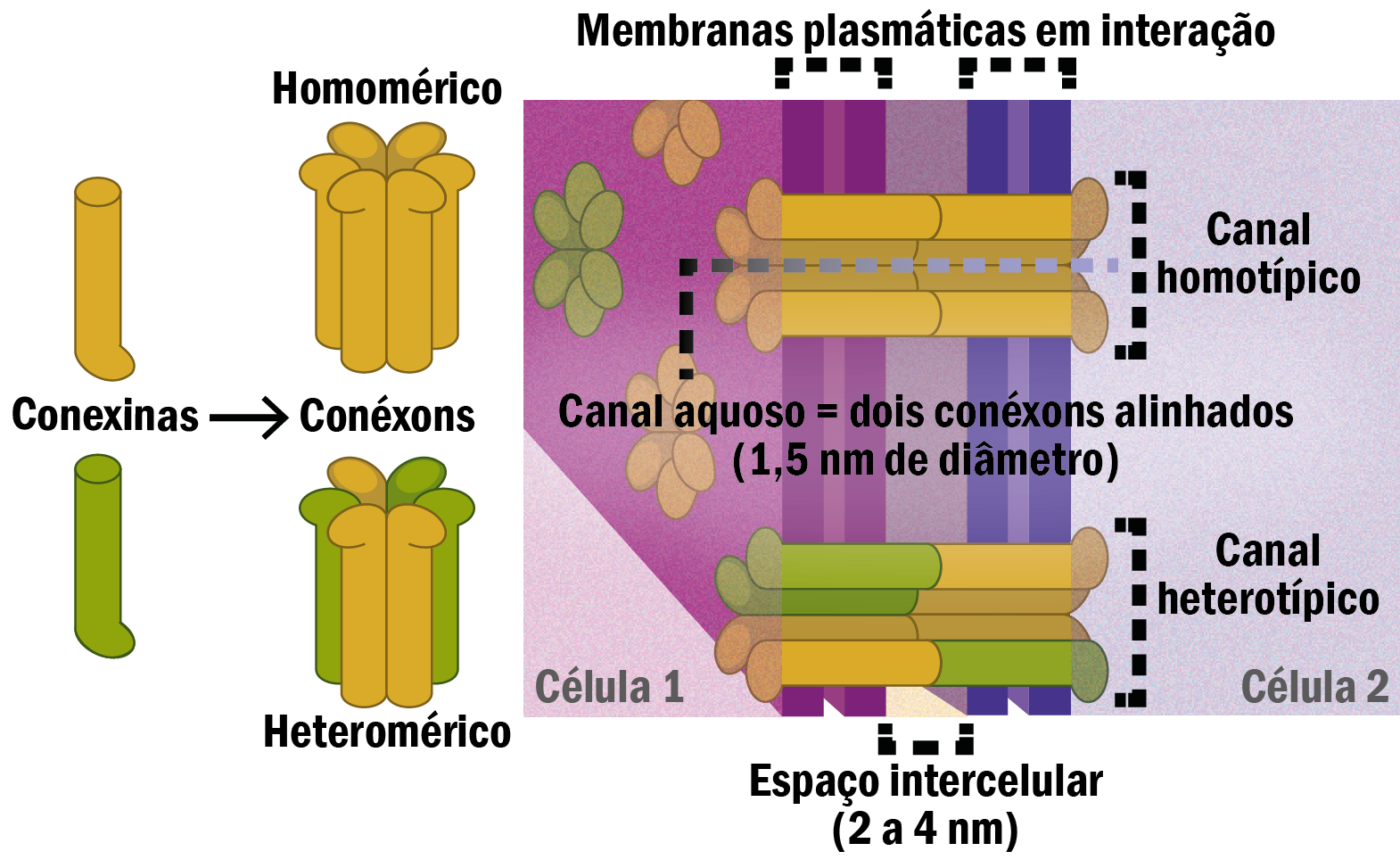
Figura 12 – (Créditos: Edilson Jacob)
As junções comunicantes são também importantes nas sinapses elétricas onde o sinal é transmitido via junções comunicantes; na embriogênese onde a distribuição de moléculas informativas ocorre através das células em migração; ou nos músculos cardíaco e liso onde ocorre um acoplamento elétrico coordenando as atividades destas células, por exemplo, movimentos peristálticos. Outro exemplo importante da função das junções comunicantes ocorre quando glicose é necessária na corrente sanguínea, ou seja, em períodos após refeição para aumentar a glicemia, o sistema nervoso estimula as células do fígado (hepatócitos) a iniciarem a quebra do glicogênio, como nem todos os hepatócitos são individualmente estimulados, o sinal é disparado de uma célula a outra através de junções comunicantes, levando a degradação do glicogênio hepático.
Junções celulares também estão presentes em invertebrados, em alguns casos as proteínas responsáveis pela junção apresentam um nome diferente, em outros a junção apresenta características específicas. Quando se trata da junção comunicante, a proteína responsável formadora do canal funcional é a proteína inexina, que desempenha a mesma função, permitindo a comunicação entre células garantindo o perfeito desenvolvimento. Para adesão célula-célula tipo junção de oclusão, a presente em invertebrados é denominada junção septada. As junções septadas formam um cinturão continuo ao redor de cada célula epitelial, e as membranas plasmáticas que interagem são ligadas por proteínas organizadas em linhas paralelas com periodicidade regular.
Interações celulares
Interação célula-célula transiente
As interações célula-célula transientes são resultado da resposta imune do organismo, que ocorre em locais de inflamação ou na recirculação de algumas células entre tecidos e corrente sanguínea. A maioria dos leucócitos circula no sangue sem interagir com as outras células sanguíneas ou com células endoteliais que revestem os vasos sanguíneos. Entretanto, os linfócitos participam de um contínuo processo de recirculação através dos tecidos linfóides.
Esse processo denominado endereçamento ou homing envolve muitas e diversas moléculas de adesão que auxiliam os linfócitos a retornar a vários compartimentos linfóides do corpo. A interação linfócitos-células endoteliais requer dois tipos de proteínas de adesão celular: selectinas e integrinas. Os leucócitos utilizam um mecanismo similar para escapar dos vasos sanguíneos de modo a atingir os sítios inflamatórios. A migração dos leucócitos da corrente sanguínea ocorre também em várias etapas (Figura 13):
- leucócitos na circulação resistem a forças tencionais de modo a diminuir a velocidade ao longo do percurso pelo endotélio vascular;
- Uma fraca adesão ao endotélio, sob condições de fluxo lento, faz com que os leucócitos rolem ao longo da superfície do endotélio, as selectinas presentes na superfície das células endoteliais ligam-se a carboidratos específicos na superfície dos leucócitos;
- As integrinas receptoras β1 e β2 para os ligantes endotelias VCAM-1 e ICAM-1 (superfamília das imunoglobulinas) são rapidamente ativadas na superfície dos leucócitos durante o rolamento, mediadores químicos do local da inflamação estimulam a ativação dessas integrinas β1 e β2 que reforçam a ligação dos leucócitos as superfícies celulares endoteliais;
- Ocorre uma forte interação e a migração transendotelial é mediada por integrinas que interagem com ligantes mas superfícies das células endoteliais.
Figura 13 – (Créditos: Edilson Jacob)
Interação célula-célula estável
As interações célula-célula estáveis são responsáveis pela organização das células em tecidos. É possível observar a importância das zônulas de oclusão em diferentes tecidos e epitélios, como exemplo:
O epitélio intestinal, um epitélio cilíndrico simples, onde se observa uma grande quantidade de células absortivas que são responsáveis pela transferência de nutrientes do lúmen do intestino para o tecido subjacente que seguem pela corrente sanguínea e vão nutrir outros tecidos. Nesse epitélio, as junções de oclusão forçam a passagem da glicose pela via transcelular, a presença de transportadores simporte que levam a internalização da glicose a partir do lúmen do intestino orientada pelo gradiente de sódio (transporte ativo dirigido por gradiente iônico), mesmo em baixas concentrações de glicose no lúmen, e na sequência essa glicose é transportada para outros tecidos subjacentes por proteínas carreadoras específicas por difusão facilitada a favor do gradiente de concentração. Dessa forma a junção de oclusão garante a polaridade celular e o transporte celular sem desperdício de energia (Figura 14).
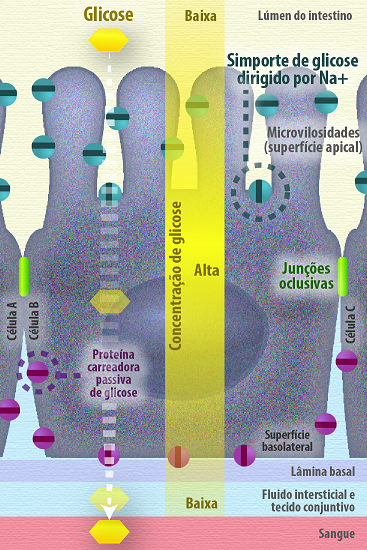
Figura 14 – (Créditos: Edilson Jacob)
Diferentes junções de oclusão selam de forma diferente, e a selagem pode não ser absoluta, embora impermeável a macromoléculas, sua permeabilidade a pequenas moléculas varia em diferentes epitélios, isso pode ser explicado pela diferente composição de proteínas que compõem as junções. No epitélio intestinal é 10.000X mais permeável a íons inorgânicos (sódio, por exemplo) do que as junções de oclusão no epitélio que reveste a bexiga, o urotélio.
A principal proteína transmembrana, essencial para formação da junção de oclusão é a proteína claudina. Camundongos que possuem mutações no gene claudina-1 não formam junções de oclusão entre as células da camada epitelial da pele, e como resultado, as crias perdem água rapidamente por evaporação pela pele e morrem em poucos dias após o nascimento.
Barreiras também importantes são formadas com as junções de oclusão, como a barreira hemato-encefálica e a barreira hemato-testicular, que garantem que microorganismos ou drogas não atravessem essa junção e causem danos nesses compartimentos. A barreira hemato-encefálica é formada pelas junções de oclusão e componentes adicionais, como: lamina basal dos capilares endoteliais e os pés terminais dos astrócitos no sistema nervoso central garantindo que moléculas cheguem ao SNC atravessando a via transcelular (Figura 15). A barreira hemato-testicular separa os compartimentos: basal e adluminal no epitélio germinativo, garantindo a espermatogênese correta sem interferência de drogas e microorganismos (Figura 16).
Figura 15 – (Créditos: Edilson Jacob)
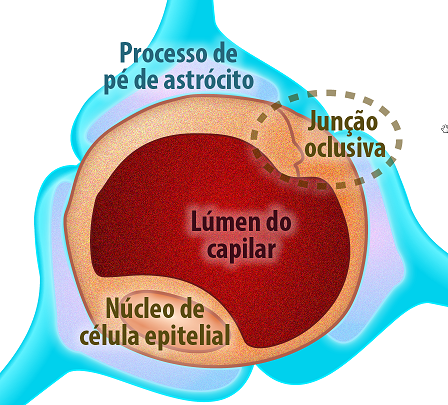
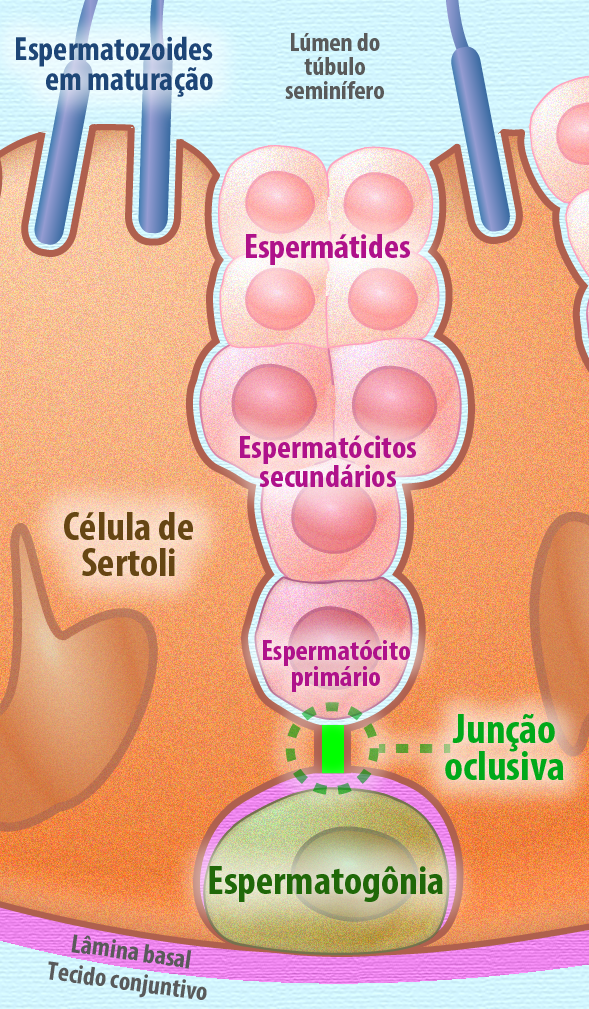
Figura 16 – (Créditos: Edilson Jacob)
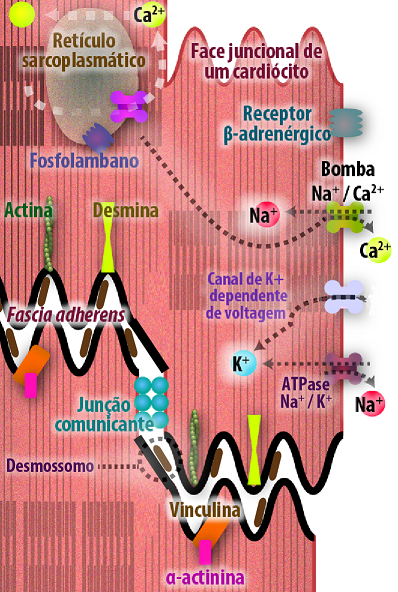
Além da junção de oclusão as junções de adesão célula-célula também desempenham funções significativamente importantes, tanto no tecido muscular cardíaco quanto na fusão entre espermatozóide e ovócito. O tecido muscular cardíaco é formado por células denominadas cardiomiócitos, que são unidos por complexos juncionais especializados, os discos intercalares. Organizados em componentes transversais e longitudinais, responsáveis principalmente pela resistência ou comunicação, respectivamente. No componente transversal encontram-se as junções de ancoragem (zônula de adesão e desmossomos), garantindo à resistência da célula as contrações. E no componente longitudinal, as junções comunicantes, são responsáveis pela rápida transmissão de sinal elétrico, levando à contração muscular sincronizada (Figura 17).
Figura 17 – (Créditos: Edilson Jacob)
Na fecundação, momento em que os núcleos dos gametas: feminino e masculino se unem para dar início ao desenvolvimento do embrião, a adesão das membranas celulares ocorre a partir do reconhecimento e interação do tipo célula-célula. Na membrana do ovócito II a presença de um complexo formado por integrina e tetraspanina reconhece e se liga a um heterodímero de fertilina alfa e beta presente na membrana celular do espermatozoide, essa interação é suficiente para levar a fusão das membranas e união dos núcleos dos gametas (Figura 18).
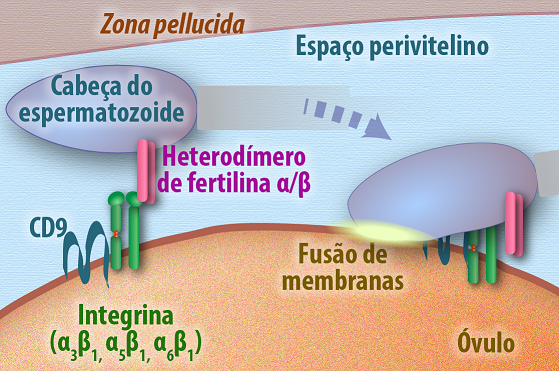
Figura 18 – (Créditos: Edilson Jacob)
As interações célula-célula executam importantes tarefas no organismo, algumas vezes bloqueiam a passagem de substâncias, outras garantem a adesão para migração celular ou integridade do tecido, permitem a comunicação celular para o bom funcionamento e desenvolvimento de um organismo. Sem dúvida, executam funções essenciais mantendo um perfeito metabolismo e organização celular (figura 19).
Figura 19 – Idem a figura 7 (Créditos: Edilson Jacob)