Arthur Tributino Menezes
Introdução
Boa parte dos seres vivos possui uma série de mecanismos e sistemas biológicos em seu corpo que lhes permite interagir com o ambiente à sua volta, se estabelecer e viver nele. Tais sistemas são bastante conhecidos em diversos organismos, como o sistema digestivo, o sistema circulatório, o sistema reprodutor, o sistema excretor, o sistema respiratório, entre outros. Esses organismos, sejam eles animais ou plantas, irão contar com esses sistemas para explorar o ambiente, encontrar alimento, se defender de predadores, digerir alimentos e absorver seus nutrientes, distribuir esses nutrientes pelo corpo, eliminar subprodutos do metabolismo, se reproduzir, entre outros.
No corpo humano isso não é diferente. Como bem sabemos, possuímos diversos órgãos que constituem os sistemas do nosso corpo e trabalham juntos para fazê-lo funcionar de maneira adequada. Se tomarmos como exemplo o sistema circulatório, identificamos o coração como órgão principal nesse sistema, que funciona como uma bomba impulsionando o sangue através dos vasos do corpo todo. Há ainda a interação do sistema circulatório com o respiratório, quando ocorre a oxigenação do sangue nos alvéolos pulmonares. Enfim, o corpo humano é um ótimo exemplo para termos noção da complexidade e organização que os sistemas biológicos podem assumir em um ser vivo.
Se olharmos esses sistemas mais de perto identificamos os diferentes órgãos que fazem parte de cada um. E se olharmos os órgãos mais de perto iremos poder explorar melhor o seu funcionamento, através da análise de seus tecidos, ou seja, dos seus conjuntos de células com funções em comum que trabalham em sintonia. Nesse texto iremos trabalhar na interface entre os níveis de organização de células e tecidos, e explorar melhor como as células se organizam para constituir um tecido biológico. Iremos perceber que a mesma colaboração entre diferentes órgãos em um sistema biológico pode também ser vista entre as células e outros componentes de um único tecido, trabalhando juntos para permitir a um determinado órgão desempenhar a sua função.
Conjuntos de células formam tecidos
Já trazemos do colegial a ideia de que células com função comum se reúnem em tecidos no corpo de um organismo vivo. Mas como será que essas células estão organizadas para constituir esses tecidos? Elas estariam lado a lado e umas sobre as outras como os blocos em uma parede? Será que elas se organizam de uma mesma forma em todos os tecidos? Será que há algum tipo de interação entre essas células, ou cada uma trabalha encerrada em si mesmo, fazendo o seu trabalho? Quais são as implicações da forma como as células se organizam em um tecido para que elas possam desempenhar as suas funções de forma adequada?
Estas perguntas servem para nos fazer olhar com mais detalhes para a organização de um tecido, e tentar entender melhor como tal organização influencia no papel que o tecido terá no órgão do qual faz parte.
Tipos de tecidos
Os tecidos do corpo humano são constituídos de células e de um material secretado por elas conhecido como matriz extracelular. Essa matriz é composta por diversos tipos de moléculas e fibras e serve para dar suporte para as células no tecido e também para várias outras funções. As células estão intimamente relacionadas com a matriz extracelular, de modo que poderíamos enxergá-las, células e matriz, como uma unidade funcional. Os elementos da matriz extracelular são constantemente produzidos e degradados pelas próprias células em um processo de remodelamento contínuo. Alguns desses elementos se conectam às células fisicamente, através de receptores nas membranas das células que por sua vez estão ligados ao seu citoesqueleto. Tal organização irá permitir diversas interações entre as células e a matriz extracelular.
Podemos distinguir quatro tipos básicos de tecidos no corpo humano: tecido epitelial, tecido conjuntivo, tecido nervoso e tecido muscular. Essa classificação leva em conta a constituição e organização de cada tipo de tecido. O tecido conjuntivo é o mais abundante em matriz extracelular. Nos tecidos epiteliais e musculares a matriz extracelular aparece como uma lâmina basal. Já no tecido nervoso ela é bem escassa.
Matriz extracelular (MEC)
Como mencionado anteriormente, a matriz extracelular, à qual podemos nos referir com a abreviação MEC, é composta de material secretado pelas próprias células do tecido do qual ela faz parte. A MEC participa da composição de diversas partes do corpo humano assumindo diferentes aspectos. Por exemplo, o globo ocular é composto por uma MEC transparente que permite a passagem da luz para podermos enxergar. Nos ossos a MEC sofre um processo de calcificação para formar uma estrutura rígida que é o próprio osso. A MEC também forma os tendões das nossas articulações.
Apesar dos exemplos discutidos acima mencionarem o corpo humano, a matriz extracelular também está presente em outros organismos. A mesogleia (um material semelhante a um gel transparente ou uma gelatina) das águas vivas é composta por matriz extracelular. O exoesqueleto de besouros e lagostas é formado pela MEC. A MEC também está relacionada com a cutícula de vermes, a concha de moluscos, a parede celular de fungos e plantas. Enfim, aqui já podemos ter uma ideia da importância e da versatilidade da MEC nos organismos vivos.
O papel da MEC não se restringe simplesmente a dar suporte para as células. As moléculas constituintes da MEC irão interagir com as células participando de processos de migração celular, proliferação, desenvolvimento, forma e função das células, entre outros.
Composição da MEC e algumas funções
Quanto a sua constituição, podemos organizar os componentes da MEC em duas categorias: fibras (colágeno e fibras elásticas) e substância fundamental (glicosaminoglicanos, proteoglicanos e glicoproteínas). (Figura 1).
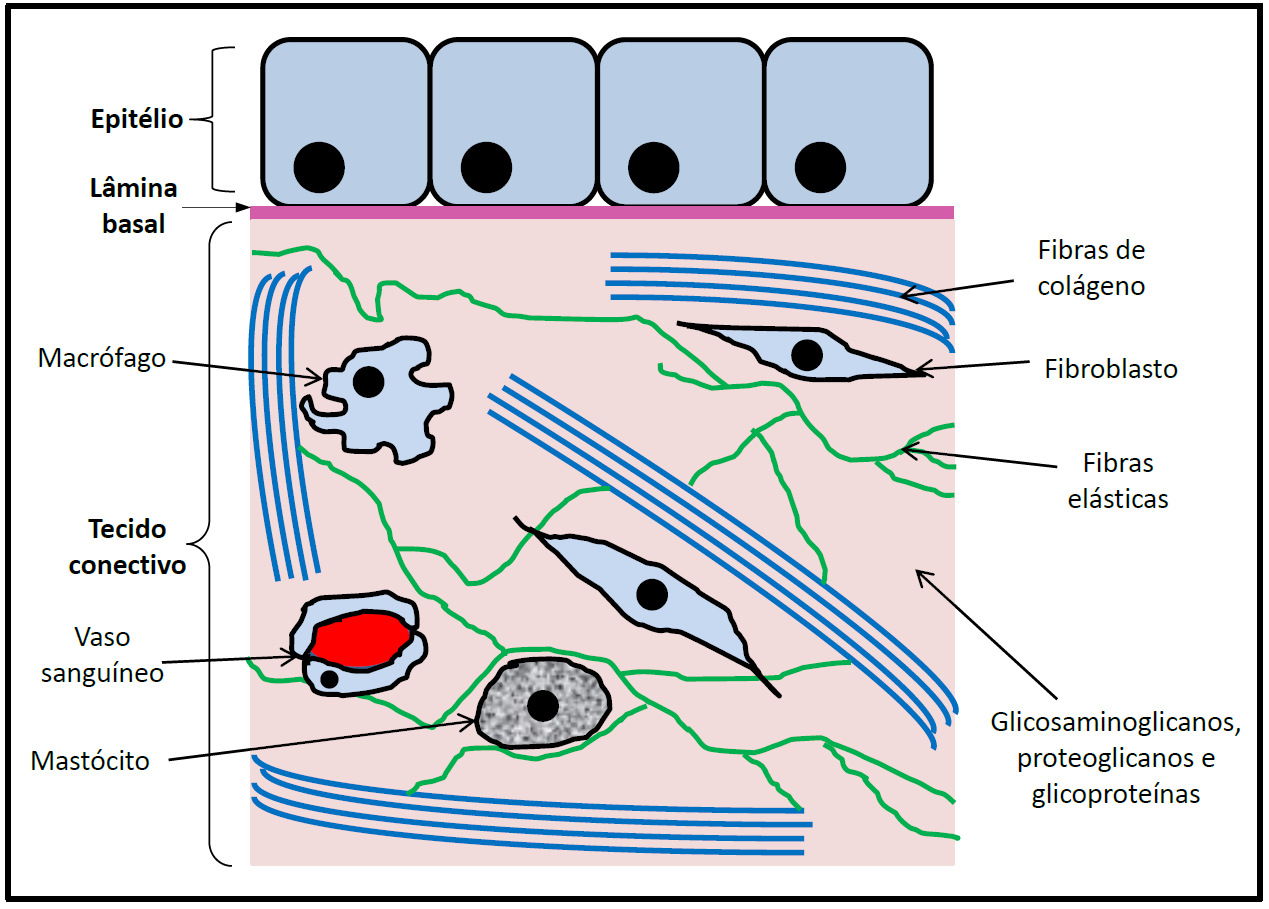
Figura 1 – Composição e organização geral da matriz extracelular (MEC). Nesse esquema está representada parte da MEC de um tecido conjuntivo subjacente a um epitélio. A lâmina basal faz a conexão entre essas duas estruturas (ver mais adiante no texto).
Créditos: Arthur Tributino Menezes
As fibras de colágeno são o componente mais abundante na MEC. Elas apresentam uma estrutura em tripla hélice que as confere grande resistência, e se apresentam em uma grande variedade de tipos com diferentes funções. As fibras colágenas irão interagir com outras moléculas na matriz, inclusive com outros tipos de fibras colágenas, e também com as células do tecido. Os fibroblastos são as principais células responsáveis pela produção dos elementos constituintes da MEC, inclusive das fibras de colágeno. Estas células produzem os precursores das fibras colágenas e orientam a sua organização no tecido. Além disso, essas células terão um papel no remodelamento do colágeno, inclusive na formação dos tendões no nosso corpo.
Além das fibras colágenas, na MEC está presente outro tipo de fibras, que são as fibras elásticas. Como o próprio nome sugere, essas fibras irão conferir elasticidade aos tecidos, o que em alguns casos é uma propriedade essencial. Tecidos como os pulmões e os vasos sanguíneos sofrem uma distensão intensa e têm a capacidade de voltarem a sua forma original, sendo esse um processo contínuo. Esses tecidos possuem essa elasticidade e maleabilidade graças às fibras elásticas. As fibras elásticas estão associadas com as fibras colágenas, e essas últimas irão limitar a distensão do tecido para evitar o seu rompimento, já que as fibras colágenas não são distensíveis como as elásticas.
Como mencionado anteriormente, além das fibras, a MEC é constituída por uma substância fundamental composta por glicosaminoglicanos (GAGs), proteoglicanos e glicoproteínas.
As GAGs são cadeias polissacarídicas compostas por várias unidades dissacarídicas que se repetem. Por serem muito rígidas e hidrofílicas, as GAGs não se dobram como outras estruturas longas (por exemplo, proteínas). Assim, elas apresentam uma estrutura que ocupa um grande volume, de modo que podem formar géis ainda que estejam em uma concentração muito baixa no meio. As GAGs, por apresentarem cargas negativas, atraem um grande número de cátions, o que leva também à atração de uma grande quantidade de moléculas de água. Isso faz com que a matriz “inche”, ficando com uma alta pressão de turgor, de modo a proporcionar ao tecido uma alta resistência à compressão. Esse é um papel antagônico ao das fibras de colágeno, que por sua vez irão proporcionar ao tecido uma resistência contra forças de distensão.
A hialuronana é o GAG mais simples. Dentre as funções que desempenha, a hialuronana cria espaços no tecido durante a sua formação embrionária para facilitar a remodelação do mesmo. Com sua capacidade de expandir amplamente a sua estrutura atraindo moléculas de água, a hialuronana participa, por exemplo, da formação das válvulas e septos no coração embrionário. Ela também atua como lubrificante nas articulações. Na sua atuação, a hialuronana depende de interações com outras moléculas da MEC, como proteínas e proteoglicanos, e também interage com proteínas na superfície da membrana plasmática das células.
Os proteoglicanos são compostos por GAGs covalentemente ligados a proteínas (Figura 2). A maior parte dos GAGs aparecem na MEC na forma de proteoglicanos, com exceção da hialuronana, que é um GAG mais simples.
Figura 2 – Estrutura geral de um proteoglicano. As moléculas de glicosaminoglicanos (GAGs) se associam com uma proteína compondo uma estrutura que tridimensionalmente lembra uma escova com cerdas ao seu redor
Créditos: Arthur Tributino Menezes
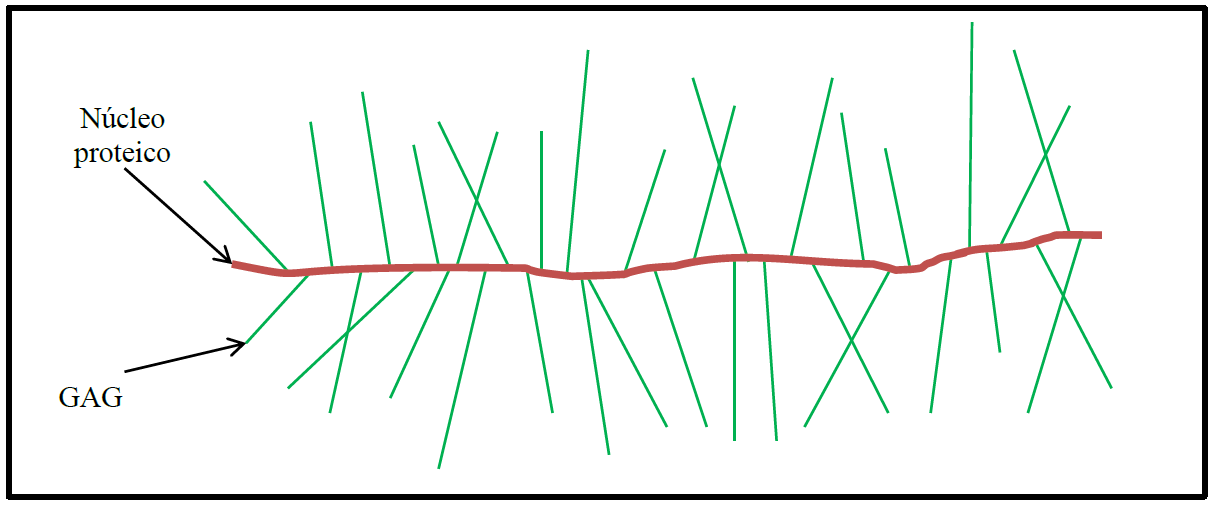
Os proteoglicanos associam-se com GAGs e outras moléculas da MEC, como as fibras de colágeno, criando uma rede bastante complexa. Por sua capacidade de formar géis de diferentes porosidades, os proteoglicanos podem atuar formando filtros seletivos para a passagem de moléculas de diferentes tamanhos, como ocorre, por exemplo, na filtragem do sangue na formação da urina nos glomérulos renais. Além disso, os proteoglicanos podem atuar na comunicação química entre as células de um tecido. Um fator de crescimento de fibroblastos, o qual sinaliza para a proliferação de diversos tipos celulares, é oligomerizado pela ação de proteoglicanos na MEC para desempenhar a sua função. Há também um papel dos proteoglicanos na resposta inflamatória, onde ele atua na imobilização de moléculas de quimiocinas, que são substâncias sinalizadoras que atraem as células de defesa do sistema imune para um determinado local para a montagem da resposta inflamatória do organismo. Essa imobilização das quimiocinas feita pelos proteoglicanos da MEC faz com que essas substâncias atuem por mais tempo localmente recrutando os leucócitos circulantes na corrente sanguínea para o local da inflamação. Os proteoglicanos irão atuar ainda na regulação da atividade de outras proteínas secretadas pelas células na MEC, inclusive de enzimas proteolíticas. Desse modo, os proteoglicanos terão um papel na síntese e degradação dos componentes da própria MEC, inclusive das fibras de colágeno. Os proteoglicanos podem ainda atuar como receptores de superfície em células, e nesses casos o seu núcleo proteico geralmente é constituído por uma proteína transmembrana componente da membrana plasmática das células.
Lâmina Basal ou membrana basal
A lâmina basal ou membrana basal é uma estrutura extracelular com um aspecto de uma fina camada de cerca de 40 a 120nm de espessura, encontrada principalmente em tecidos epiteliais, mas também aparece circundando individualmente as células musculares, os adipócitos e as células de Schwann (formadoras da bainha de mielina em alguns tipos de neurônios). A lâmina basal é formada por MEC, e, portanto irá apresentar os mesmos componentes que vimos até aqui, com algumas particularidades. A presença de lâmina basal é uma característica comum a todos os organismos multicelulares, tal é a sua importância na constituição e arquitetura dos tecidos nesses organismos.
Em geral a lâmina basal localiza-se na base dos epitélios, separando-os da matriz extracelular do tecido conjuntivo subjacente. Ela tem uma função importante na manutenção da adesão entre a epiderme (a pele) e o tecido conjuntivo subjacente, de modo que quando essa função está prejudicada o indivíduo pode desenvolver uma doença com consequências graves (esse assunto será melhor discutido mais adiante). Além disso, a lâmina basal pode estar presente entre duas camadas celulares, apresentando assim um papel de filtragem de substâncias (Figura 3).
Dentre as demais funções da lâmina basal estão: determinar a polaridade celular, influenciar o metabolismo celular, organizar as proteínas nas membranas plasmáticas em células adjacentes, promover a sobrevivência, a proliferação e a diferenciação celular, atuando ainda como via para a migração celular.
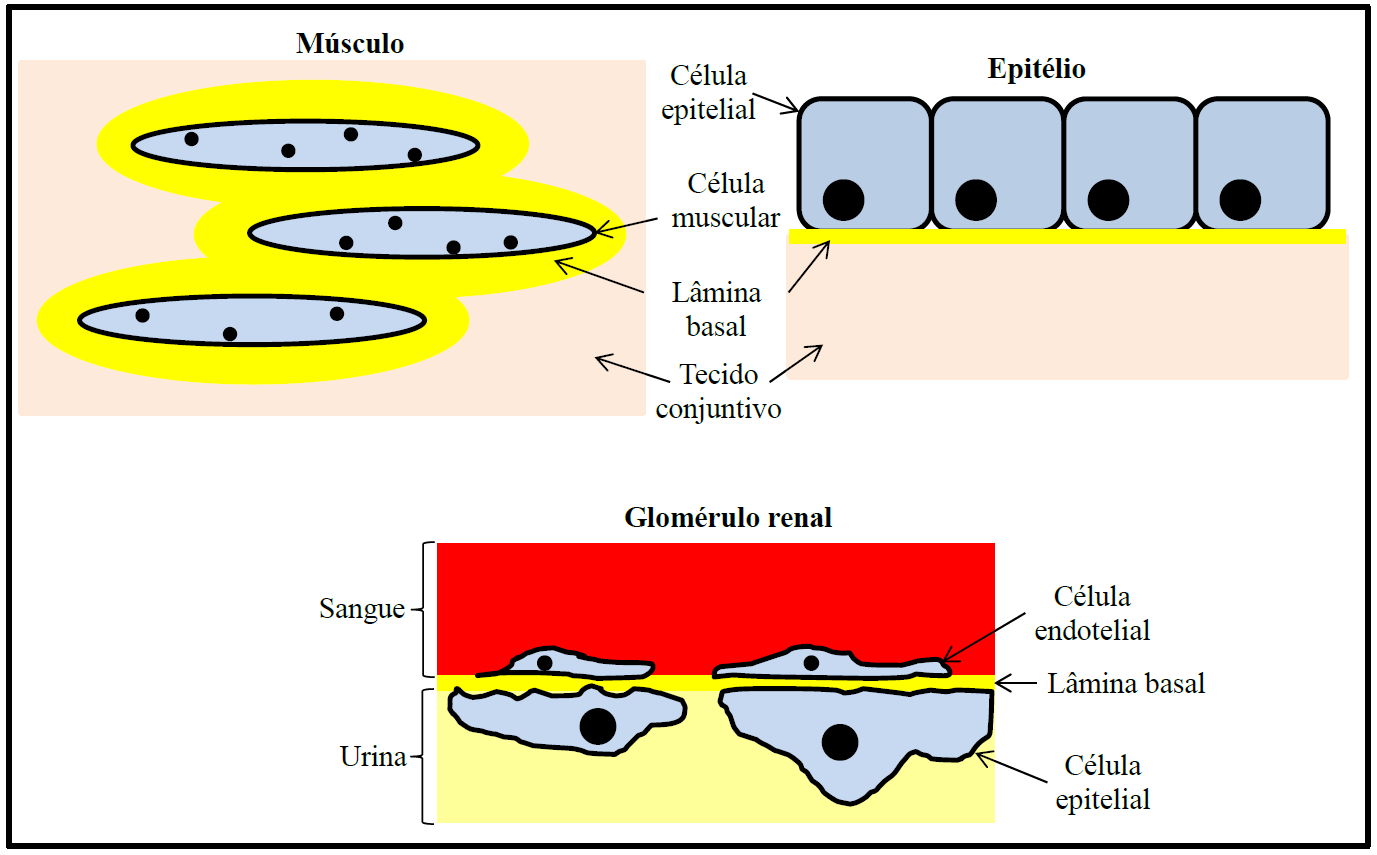
Figura 3 – Diferentes tecidos onde a lâmina basal pode ser encontrada. A função da lâmina basal está relacionada com o tipo de tecido onde ela se encontra e também com a sua localização nesses tecidos.
Créditos: Arthur Tributino Menezes
A lâmina basal é construída a partir da secreção de moléculas das células ao seu redor. Por exemplo, a lâmina basal de um epitélio irá receber componentes tanto das células epiteliais na parte de cima quanto do tecido conectivo subjacente. Em sua constituição ela irá apresentar os GAGs, proteoglicanos, glicoproteínas e fibras que vimos na composição da MEC anteriormente. Porém, o principal constituinte da lâmina basal é uma glicoproteína que recebe o nome de laminina. A laminina é o componente mais abundante em lâminas basais em início de formação, e acredita-se que elas sejam as organizadoras primárias da estrutura em forma de camadas. Além da laminina, os principais componentes da lâmina basal são o colágeno tipo IV, o nidogênio e a perlecana. Na formação da lâmina basal, as moléculas de laminina interagem entre si e com proteínas na membrana das células formando uma rede no espaço extracelular que constituirá um arcabouço para a organização das demais moléculas constituintes da lâmina basal.
Como já mencionado, a lâmina basal tem um papel importante na formação da urina nos glomérulos renais. Mutações que afetem os componentes da lâmina basal nesse tecido podem levar um indivíduo a desenvolver doenças como a Síndrome de Alpert (ver mais adiante). Esse papel seletivo da lâmina basal não está restrito aos rins. Ela também irá atuar de forma seletiva na regulação dos tipos de células aos quais ela permitirá que a atravessem. Os fibroblastos dos tecidos conectivos, por exemplo, em geral não são capazes de atravessar a lâmina basal para chegar à epiderme. Por outro lado, os leucócitos que são recrutados durante uma resposta inflamatória têm passagem garantida para poderem desempenhar o seu papel na defesa do organismo. A lâmina basal também tem um papel importante na cicatrização de feridas. Quando ocorre uma lesão, a lâmina basal normalmente é preservada, e assim fornece um suporte para a migração das células na cicatrização de modo que o tecido se reconstitua de maneira mais fácil.
Nas junções neuromusculares, aquelas formadas entre um neurônio motor e uma célula muscular, a lâmina basal tem um papel bastante interessante. Nessa estrutura a lâmina basal se localiza externamente à membrana muscular na região de sinapse entre o nervo e o músculo. Experimentos mostraram que a lâmina basal tem um papel fundamental na reconstituição da sinapse após algum tipo de lesão que afete o nervo ou o músculo. No caso de a célula muscular se degenerar, a lâmina basal permanece intacta, e um neurônio pode novamente formar a sinapse com a região da lâmina basal onde antes havia uma sinapse. Assim, a lâmina basal por si só é capaz de guiar a regeneração dos terminais dos nervos motores. Em outra situação, se somente a célula muscular se regenera após a lesão, mas não o neurônio, os novos receptores de acetilcolina produzidos pela célula muscular irão se localizar na fenda sináptica nas mesmas posições onde estavam os receptores antigos antes da lesão. Assim, a lâmina basal é capaz de guiar a regeneração, na junção neuromuscular, de componentes tanto da célula muscular quanto do neurônio.
Outro ponto que vale a pena explorar um pouco é a importância da ligação das células à MEC. Como já vimos, a matriz extracelular faz muito mais do que apenas servir de suporte mecânico para as células. Estudos demonstraram que alguns tipos de células apresentam uma sobrevivência dependente de adesão à MEC, ou seja, estas células entram em apoptose caso sejam impedidas de se ligar à MEC. A ligação das células à MEC se dá por vários mecanismos, sendo um deles através de proteínas de adesão. As integrinas são proteínas transmembrana de adesão relacionadas com a ligação das células à lâmina basal. Essa ligação faz mais do que simplesmente manter as células unidas à MEC, pois as integrinas também participam de sinalizações de vias metabólicas intracelulares. Elas irão mediar, por exemplo, a sinalização da ligação das células à MEC, de modo que a célula receba essa informação e “perceba” que está ligada à MEC, o que impede o início do programa de morte celular. Além de influenciar na sobrevivência das células, a MEC também irá interferir na proliferação celular.
Relevância Clínica
Doenças relacionadas à MEC
Síndrome de Eherles-Danlos tipo VII
A Síndrome de Eherles-Danlos apresenta-se em diferentes formas, sendo distinguidos dez tipos dessa doença. Trata-se de uma doença com fatores hereditários, apresentando na maioria dos casos um padrão de herança autossômico dominante. Algumas das mutações relacionadas a essa doença afetam genes relacionados com a síntese de diferentes tipos de colágeno. Como vimos anteriormente, o colágeno é a proteína mais abundante na matriz extracelular, de modo que tais alterações irão afetar principalmente os tecidos conjuntivos. De um modo geral, os sintomas mais comuns dessa síndrome são a hiperextensibilidade da pele, a hipermobilidade articular, entre outros. Os indivíduos portadores dessa síndrome acabam morrendo muitas vezes pelo rompimento de grandes vasos sanguíneos, como a aorta.
Escorbuto
O escorbuto é uma doença relacionada à falta de vitamina C. Seus sintomas incluem sangramentos nas gengivas, perda dos dentes, dores nas articulações, entre outros. Antigamente, quando não se conhecia a causa do escorbuto, muitos marinheiros morriam dessa doença. Como os marinheiros passavam grandes períodos de tempo em viagem pelos mares eles acabavam tendo uma dieta pobre em vitamina C. Um dos motivos era o fato de que as frutas, que seriam uma fonte principal de vitamina C, não eram alimentos apropriados para serem estocadas por muito tempo por estragarem rápido. Sendo assim, ainda que naquela época as pessoas não sabiam disso, a falta de comer frutas levava os marinheiros a desenvolver essa doença.
A vitamina C é um importante cofator na via de síntese do colágeno, de modo que a sua deficiência leva à produção de fibras colágenas defeituosas, o que pode levar uma pessoa a desenvolver o escorbuto (ver tópico sobre Síntese do Colágeno para saber mais).
Osteogênese imperfeita
Como vimos anteriormente, a MEC participa da formação dos ossos do corpo humano. Ela vai sofrendo um processo de deposição de cálcio para dar origem à estrutura rígida de sustentação do nosso corpo que são os ossos do nosso esqueleto. Sendo assim, é de se esperar que problemas que afetem os componentes dessa MEC acabem por interferir na formação dos ossos, e é exatamente isso que ocorre na osteogênese imperfeita. Essa doença possui um caráter hereditário, e a sua causa está relacionada com defeitos nos genes que codificam a síntese de colágeno. Dessa forma, os ossos do indivíduo portador dessa doença têm uma má formação, de modo que ficam bastante frágeis, daí a doença também ser conhecida popularmente como “ossos de vidro”. O indivíduo que nasce com essa doença pode já sofrer fraturas durante o seu nascimento, ao longo do trabalho de parto.
Epidermólise bolhosa
Esta doença caracteriza-se pela formação de bolhas na pele causadas por traumas leves, dentre outras complicações mais graves que podem afetar outros tecidos, como partes do intestino. É uma doença hereditária causada por alterações em genes que codificam componentes relacionados à lâmina basal, como laminina, integrina e colágeno. Essas alterações interferem na junção entre tecido conjuntivo e epitélio, culminando no aparecimento dos sintomas citados.
Síndrome de Alport
Os portadores dessa doença apresentam problemas renais, também já tendo sido relatados casos de surdez neurossensorial e alterações oculares. É uma doença genética, relacionada com defeitos no gene que codifica um tipo de colágeno presente na lâmina basal de diversos tecidos, como nos glomérulos renais. Por esse motivo, a filtragem do sangue nos rins não é feita de maneira correta, e o portador dessa doença acaba por perder nutrientes em excesso na formação da urina, bem como não eliminar corretamente os metabólitos de seu organismo.
Links interessantes
- Vídeo sobre a composição da Matriz extracelular (em espanhol):
- Vídeo-aula do canal da Khan academy sobre Matriz extracelular:
Referências
Livros:
- Alberts, et al. 2011. Biologia Molecular da Célula. 5ª ed. Artmed;
- Junqueira, L.C.U. & Carneiro, J.. 2008. Histologia Básica. 11ª Ed. Rio de Janeiro: Guanabara Koogan.
Sites:
- Associação Brasileira de Síndrome de Ehlers-Danlos e Hipermobilidade. Disponível em: <https://sedbrasil.wixsite.com/home>.
Acesso em: 02/03/2022. - Osteogênese imperfeita. Disponível em: <https://www.hcrp.fmrp.usp.br/sitehc/informacao.aspx?id=549&ref=1&refv=12>.
Acesso em: 06/11/2016. - Síndrome de Alport: Como ela é?. Disponível em: <https://www.abc.med.br/p/sinais.-sintomas-e-doencas/814729/sindrome+de+alport+como+ela+e.htm>.
Acesso em: 06/11/2016.
Artigos:
- Almeida Jr, Hiram Larangeira de. Genética Molecular das Epidermólises Bolhosas. An. Bras. Dermatol.Rio de Janeiro,v. 77, n. 5, p. 519-532,Oct.2002. Disponível em: <https://www.scielo.br/scielo.php?script=sci_arttext&pid=S0365-05962002000500002&lng=en&nrm=iso>.
Acesso em: 06/11/2016. - Gurtler, Thaiz Gava Rigoni; Diniz, Lucia Martins; Souza Filho, João Basilio de. Epidermólise bolhosa distrófica recessiva mitis: relato de caso clínico. An. Bras. Dermatol.,Rio de Janeiro ,v. 80, n. 5, p. 503-508,Oct.2005 . Disponível em: <https://www.scielo.br/scielo.php?script=sci_arttext&pid=S0365-05962005000600009&lng=en&nrm=iso>.
Acesso em: 06/11/2016. - Leal, Fernando Antônio de Macedo et al . Síndrome de Alport: estudo de uma família. Arq. Bras. Oftalmol.,São Paulo,v. 63, n. 6, p. 455-458,Dec.2000 . Disponível em: <https://www.scielo.br/scielo.php?script=sci_arttext&pid=S0004-27492000000600005&lng=en&nrm=iso>.
Acesso em: 06/11/2016. - Nomura, Marcelo Luís; Surita, Fernanda Garanhani De Castro; Parpinelli, Mary Angela. Síndrome de Ehlers-Danlos e gravidez: relato de caso. Rev. Bras. Ginecol. Obstet.,Rio de Janeiro ,v. 25, n. 10, p. 745-748,Dec.2003 . Disponível em: <https://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-72032003001000008&lng=en&nrm=iso>.
Acesso em: 06/11/2016. - Hynes, R.O.; (2014); “Stretching the boundaries of extracellular matrix research.”; Nat Rev Mol Cell Biol 15: 761–763. Disponível em: <https://www.nature.com/nrm/journal/v15/n12/full/nrm3908.html>.
Acesso em: 06/11/2016.
RFS