Dr. Rafael Seigi Kohasigawa
Introdução
A membrana celular (tanto a plasmática quanto a das organelas) é composta por uma bicamada lipídica e proteínas associadas a ela, que podem possuir variados fins. São funções da membrana:
| Função | Descrição |
|---|---|
| Definir os limites da célula e delimitar compartimentos internos | A formação de diferentes compartimentos e a separação entre citosol e meio externo podem ser utilizadas para manter um potencial eletroquímico, energia que é útil para transportar substâncias (cotransportador Na-glicose), transmitir informações (potenciais de ação), por exemplo. |
| Organizar reações complexas | As reações metabólicas precisam de ambientes com condições específicas para ocorrer, como as que são catalisadas por enzimas lisossômicas. |
| Organizar o fluxo de informação | Possibilitar a comunicação entre células (potenciais de ação, receptores, junções comunicantes, etc.). |
| Permitir a passagem seletiva de moléculas | Moléculas hidrofóbicas passam pela bicamada lipídica, pequenas moléculas hidrofílicas podem passar por ela, moléculas maiores e íons necessitam de proteínas para fazer esse transporte. |
Tabela 1 – Funções da Membrana Plasmática.
A bicamada lipídica
Natureza química
Todos os lipídeos (ou lipídios, se preferir) da bicamada têm caráter anfifílico (parte da molécula é solúvel em meio apolar e a outra, em polar). A base estrutural dela são os fosfolipídios, mas há outras moléculas em menor quantidade, como colesterol e glicolipídeos.
Fosfolipídios
Sua estrutura é formada por uma “cabeça” polar e duas “caudas” apolares, que em animais, plantas e bactérias são predominantemente duas longas cadeias carbônicas, uma delas, em geral, com uma insaturação em cis, o que causa curvatura da cadeia.
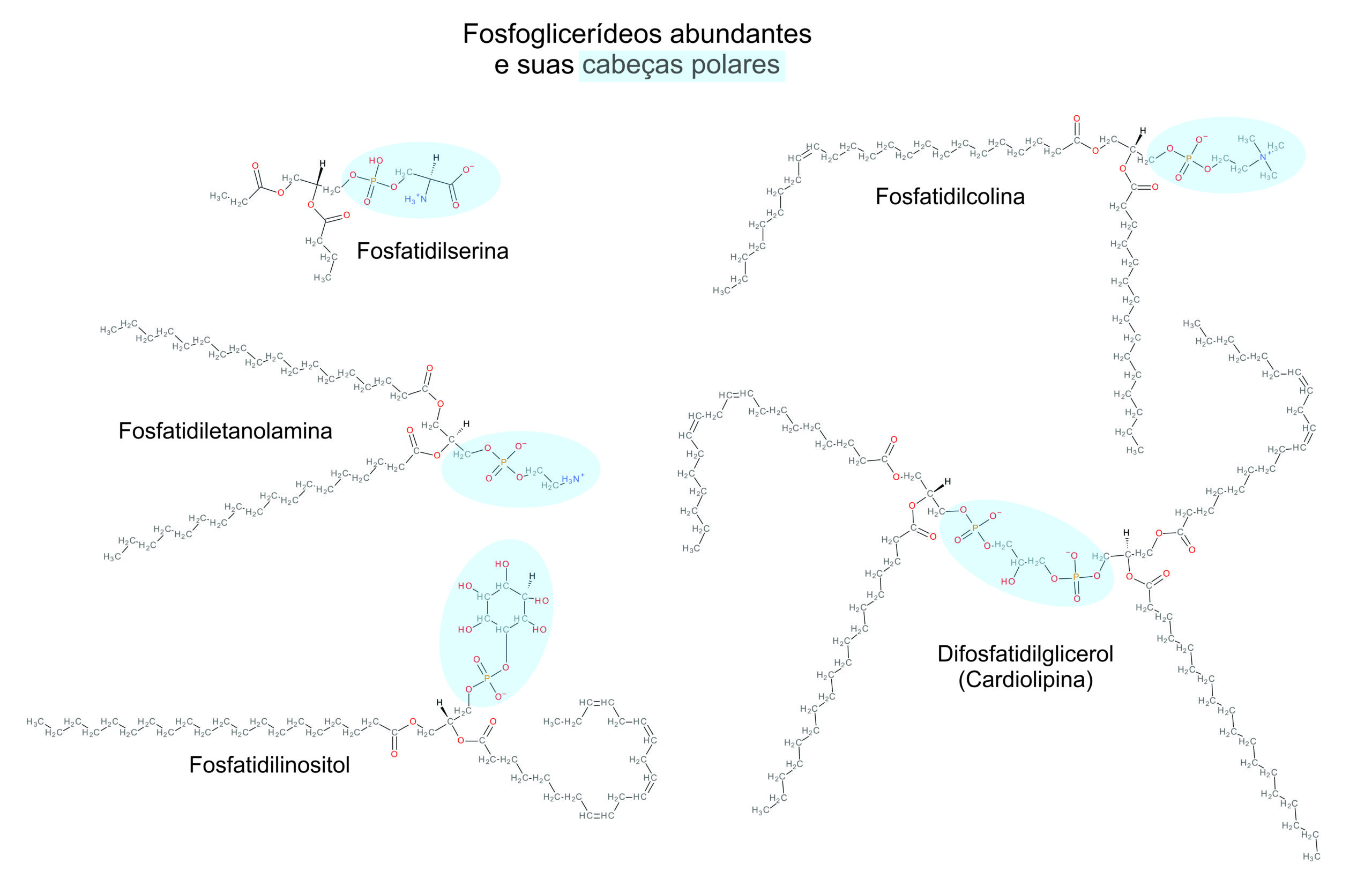
Figura 1 – Diversidade e estrutura de fosfolipídios.
Fosfoglicerídeos
São os fosfolipídeos mais abundantes. Eles apresentam um grupo fosfato, em que podem ser adicionadas outras moléculas, como aminoácidos e até outros fosfolipídios, e duas cadeias de ácidos graxos ligados cada um a um dos carbonos do glicerol por uma ligação éster.
Relevância Clínica
Um fosfoglicerídeo importante para a coagulação é o fator ativador de plaquetas, que está presente na membrana de basófilos (um tipo de leucócito) e estimula a agregação e secreção de serotonina (vasoconstritor) por plaquetas. Esse sinalizador molecular também tem efeito em vários órgãos e é importante para as respostas inflamatória e alérgica.
Esfingomielinas
São outro tipo importante de fosfolipídio, formadas por uma esfingosina ao invés de um glicerol. A esfingosina apresenta uma longa cadeia de hidrocarboneto saturado, e uma extremidade polar em que há um grupo amina, que será usado para ligar outra cadeia carbônica, e dois grupos hidroxila, um ficará livre e no outro é adicionado uma fosfocolina.

Figura 2 – Esfingomielina.
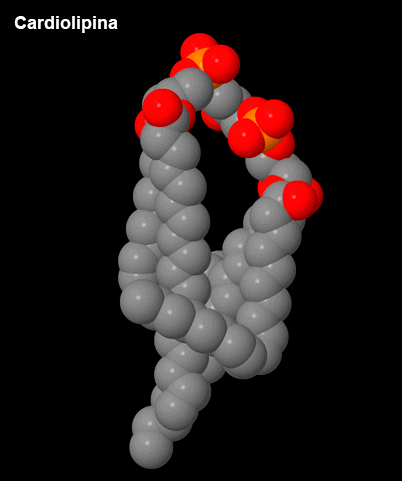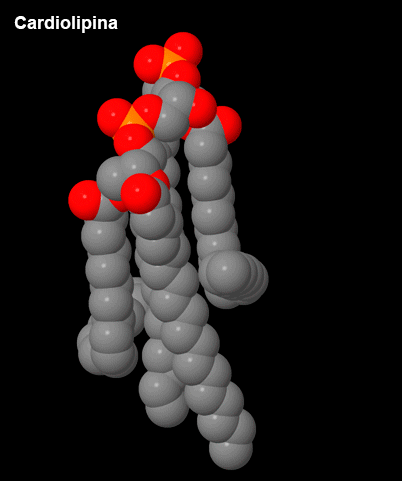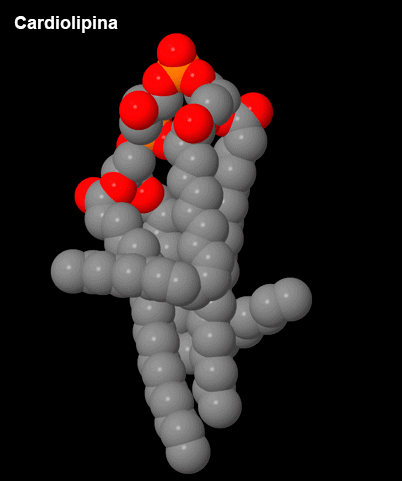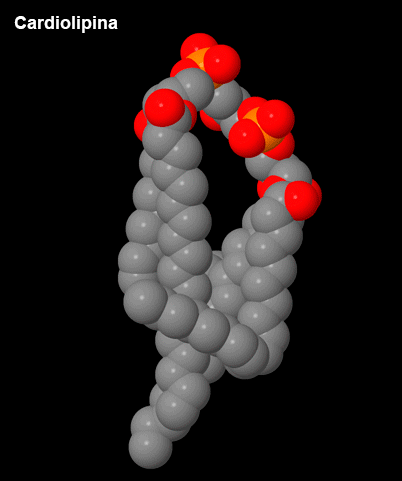
Figura 3 – Cardiolipina.
Glicolipídeos
A estrutura deles é semelhante à dos fosfolipídios, mas possuem monossacarídeos ou oligossacarídeos ao invés do grupo fosfato e, em animais, apenas a esfingosina forma glicolipídeos. Estão presentes em baixas quantidades na membrana (em geral, constituem 5% dos lipídeos da camada externa) e acredita-se que sejam importantes no reconhecimento da célula, proteção mecânica da membrana, alteração do potencial de membrana, embora nenhuma grande alteração tenha sido observada em camundongos que apresentavam deficiência em todos os glicolipídeos.
Colesterol
Molécula esteroide, possui estrutura rígida e apenas uma hidroxila. Ocupa espaços entre fosfolipídeos (há pequenos espaços devido a insaturações nas cadeias lipídicas) e é importante para a bicamada lipídica por mudar suas propriedades físicas, como fluidez, permeabilidade e flexibilidade.
Relevância Clínica
Sabe-se ao menos que os glicolipídeos possibilitam que toxinas bacterianas entrem na célula, como a toxina do vibrião colérico. Essa toxina entra apenas nas células em que há GM1 (um tipo de glicolipídeo), causando aumento de cAMP (AMP cíclico), o que leva à excreção de Na+ na luz intestinal e, consequentemente, de água também.
Figura 4 – Colesterol.
Organização estrutural da bicamada
Os lipídeos se estruturam espontaneamente em bicamadas ou micelas, pois a interação eletrostáticas das “caudas” apolares com a água é termodinamicamente desfavorável.
Essas interações não favoráveis forçam as moléculas de água próximas ao lipídeo a se arranjarem de uma forma mais organizada entre elas e envolver a molécula hidrofóbica em uma “jaula”. Esse aumento de organização eleva a energia livre, já que diminui a entropia do sistema (o sistema se tornou mais organizado devido à interação entre água e lipídeos). A energia livre pode ser atenuada com o agrupamento das moléculas apolares (ou das partes que são apolares), pois isso diminui o número de moléculas de água que envolvem os lipídeos.
Assim, fosfolipídios formam bicamadas espontaneamente em meio aquoso por dois motivos: diferença de afinidade eletrostática, pois as extremidades polares interagem melhor com a água do que com a parte hidrofóbica, enquanto as “caudas” apolares são afastadas da água, e estrutura do fosfolipídeo, que é “cilíndrica”, e não “cônica”, favorecendo a formação de uma estrutura mais “plana”.
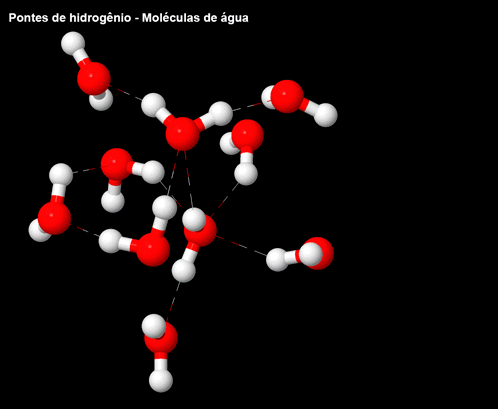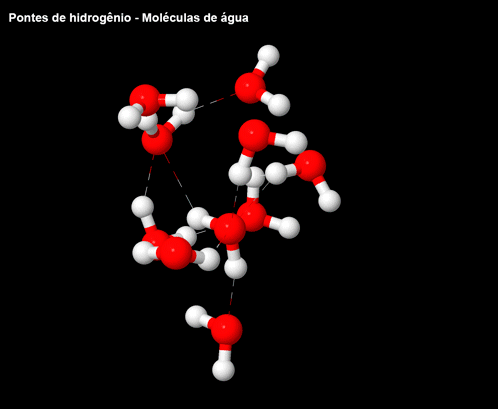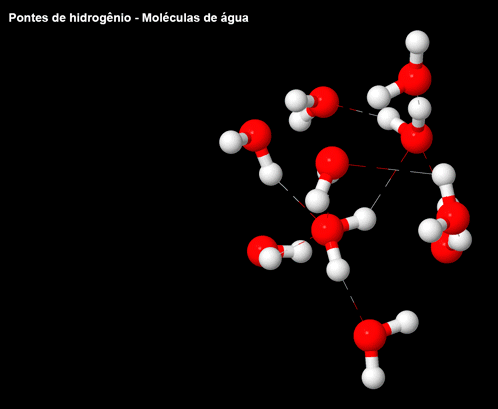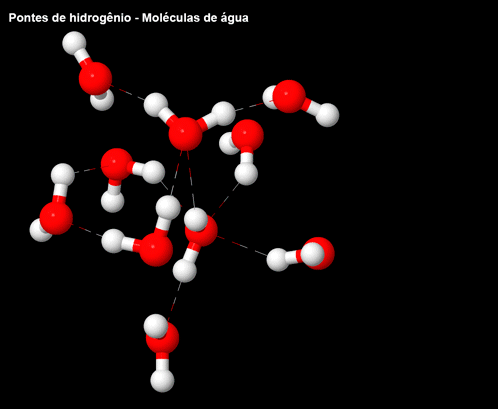
Figura 5 – Ligações/pontes de hidrogênio nas moléculas de água.
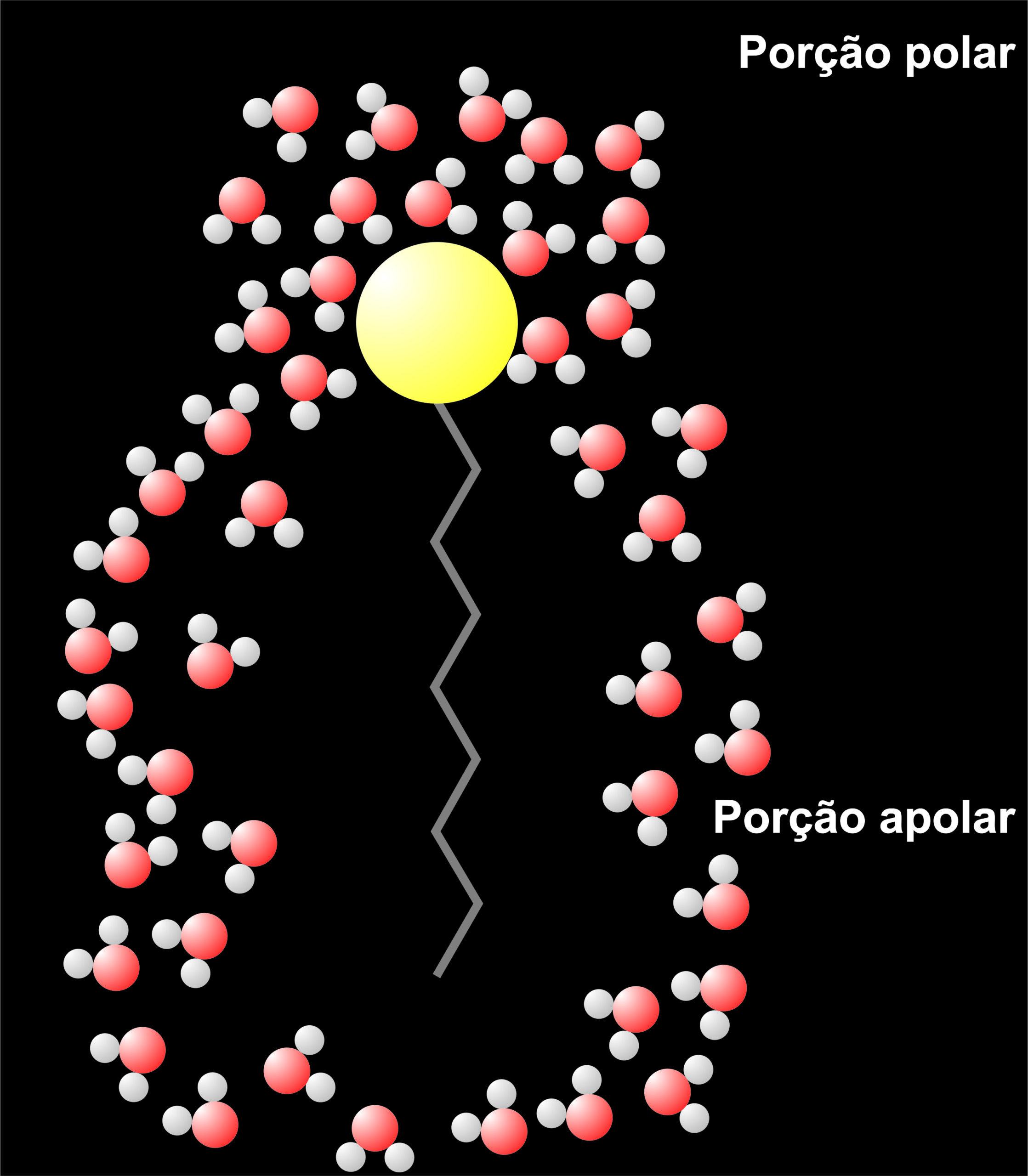
Figura 6 – Interações entre moléculas de água e fosfolipídio.
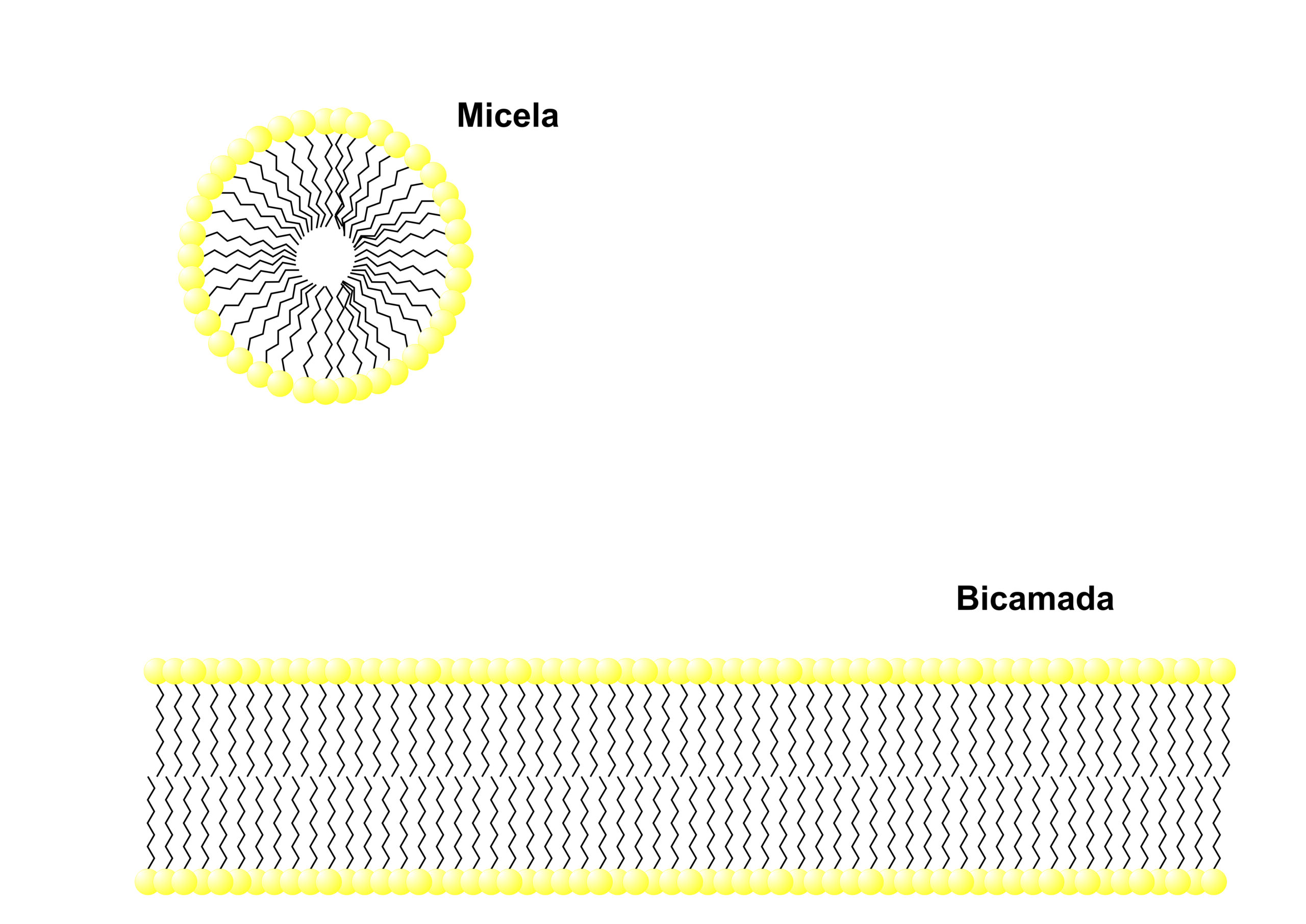
Figura 7 – Estrutura de micelas.
Essas características da bicamada fazem com que ela forme compartimentos fechados (já que esse é o único modo de uma bicamada não expor a parte apolar ao meio aquoso) e lhe dá fluidez, já que a interação entre os fosfolipídios, que se dá por forças de Van der Walls, é baixa. Ambas as características são muito importantes para a existência de uma célula, pois permite a compartimentalização celular e a difusão de lipídeos e proteínas pela membrana.
Entretanto, a diferença de afinidade eletrostática também dificulta que os lipídeos passem de um lado da membrana para o outro, o que é conhecido como flip-flop e ocorre com uma molécula em média uma vez por mês, o que é um problema para a síntese da camada externa, pois os lipídeos só são sintetizados no citosol (camada externa do retículo endoplasmático). Isso é contornado com enzimas translocadoras de fosfolipídeos, ou flipases, que fazem a rápida troca de uma camada para a outra.
Temperatura e características dos fosfolipídeos, como comprimento das cadeias e duplas ligações, alteram propriedades físicas da bicamada. De acordo com a temperatura, ela pode se tornar mais ou menos fluida, diminuindo ou aumentando a interação entre os fosfolipídios, que podem se cristalizar em temperaturas muito baixas. Cadeias curtas diminuem a tendência de interação “intracamada” e “intercamadas” dos fosfolipídios, enquanto duplas ligações dificultam a compactação dos fosfolipídios. Essas características afetam a temperatura de cristalização e a permeabilidade da bicamada, já que alteram a viscosidade dela também.
Já o colesterol diminui a permeabilidade da bicamada, mas não sua fluidez. Os anéis esteroides dele interagem com os carbonos iniciais dos fosfolipídios mais próximos, o que diminui a mobilidade deles e os agrupa mais, logo, diminui também a permeabilidade da bicamada a pequenas moléculas hidrossolúveis. Contudo, não há diminuição da fluidez, há aumento, na verdade, pois membranas ricas em colesterol tendem a não cristalizar em baixas temperaturas, já que o colesterol também diminui a interação dos fosfolipídios entre si.
Especializações da bicamada lipídica
A membrana pode possuir diversos tipos de especializações de acordo com sua localização na célula e do fenótipo celular, mas a maioria delas é devida a proteínas, então serão discutidas mais adiante neste capítulo.
Um tipo de especialização que ocorre na bicamada são as barcas lipídicas. Ao contrário do que seria esperado de uma membrana fluida, os lipídeos dela podem se organizar temporariamente em agrupamentos chamados de barcas lipídicas. Elas são formadas por lipídeos mais longos e de cadeia insaturada (com alta porcentagem de esfingofosfolipídeos) e colesterol, o que faz com que as barcas sejam mais densas do que o resto da membrana. Assim, as barcas lipídicas podem organizar proteínas de membrana, concentrando-as para serem transportadas em vesículas, para transdução de sinal (como ocorre com o receptor para insulina).
Entretanto, isso só ocorre se houver sinalização para a união desses domínios de membrana, pois eles são normalmente muito pequenos e contêm poucas proteínas.
Assimetria das camadas
Há grande diferença entre cada camada. Em geral, a cabeça dos fosfolipídios da face P apresentam carga negativa e há mais proteínas aderidas a essa face, enquanto a cabeça dos fosfolipídios e proteínas externos apresentam oligossacarídeos.
A diferença de composição gera certa diferença de potencial entre a camada externa e interna, o que terá importância para o potencial eletroquímico da membrana. Essa diferença de composição também é utilizada por animais para identificar células vivas e mortas durante eventos do desenvolvimento, como apoptose e metamorfose. Fosfatidilserina está presente na camada interna, mas quando a célula sofre apoptose ela é transferida para a camada externa, o que serve como sinal para as células próximas, como macrófagos. Isso pode ocorrer por dois mecanismos: a fosfolipídio translocase que mantém esse fosfolipídio na face P é inativada, ou uma flipase não específica é ativada e leva a fosfatidilcolina para a camada externa.
Proteínas de membrana
As proteínas de membrana dão caráter diferente a cada membrana dos variados fenótipos celulares, sendo as grandes responsáveis pelos diversos processos que ocorrem na membrana.
Elas podem ser receptores, enzimas, transportadores (poros, canais ou trocadores), “âncoras” para outras proteínas, estruturas para adesão entre células, etc.
Associação com a membrana
As proteínas se associam com a membrana dependendo de sua solubilidade:
- Proteínas de caráter anfifílico estarão parcialmente imersas na bicamada, atravessando-a (proteínas transmembrana), ou em apenas uma das camadas, pois são solúveis tanto em meio hidrofóbico como hidrofílico. A região hidrofóbica pode conter uma ou mais a-hélices ou ser composta por uma rígida estrutura de folhas-ß (conformação que se dá o nome de barril ß e é muito comum em bactérias, mitocôndrias e cloroplastos, mas não tanto em humanos);
- Proteínas solúveis em meio aquoso podem se associar à membrana por diversos tipos de âncoras lipídicas, como palmitoil, miristoil, farnesil ou glicosil fosfatidil inositol (GPI), ou por meio de outras proteínas.
As proteínas podem se associar à membrana mais fraca ou fortemente. As que se desprendem da membrana por mudanças de pH ou de concentração de outros íons são chamadas de proteínas periféricas de membrana, já as que não podem ser extraídas desse modo, mas apenas pela ação de agentes capazes de interferir na hidrofobicidade da membrana (como detergentes) são proteínas integrais de membrana.
Especializações da membrana
Assim como ocorre com lipídeos, as proteínas podem difundir-se lateralmente pela membrana e dificilmente fazem flip-flop. Então é necessário haver mecanismos para confinar proteínas específicas em determinadas regiões da membrana. Isso pode ser feito por:
- Agregação de proteínas, formando grandes complexos proteicos, o que torna o movimento delas muito mais lento se comparado com as proteínas separadas;
- Estruturas proteicas intercelulares, como as presentes em células epiteliais e que são chamadas de junções de adesão (embora essas estruturas tenham que ser mantidas em seu lugar para não realizar movimento lateral);
- Associação com macromoléculas, que podem ser intra (proteínas do citoesqueleto) ou extracelulares e uso de proteínas filamentosas para formar barreiras, criando domínios temporários ou permanentes.
Graças ao confinamento das proteínas, regiões específicas numa membrana podem existir, como o que ocorre em células epiteliais especializadas em absorção. Essas células possuem uma região apical, onde há poros, canais e trocadores, o que lhe dá a capacidade de absorção, e uma região basolateral, em que há desmossomos e junções oclusivas que permitem a união entre as células do epitélio e que não haja passagens de algumas substâncias no espaço entre as membranas plasmáticas de células adjacentes.
Vídeo ilustrando epitélio de revestimento da luz do intestino delgado: